Relationships between seedling height and nodes
Updated November 2023 to include results from 2022 season
Seedling height is positively correlated with the number of nodes, or internodes, produced along the leading shoot. If this relationship is robust it can be used to predict the number of nodes, hence the onset of the adult stage of development, on the basis of measured seedling heights.
Measurements at Ystwyth Valley Apple Breeders
To characterise this relationship under our outdoor growing conditions, we are measuring seedling height (cm) and counting the number of leaf nodes along the single leading shoot at the end of each growing season. We are treating all seedlings from a given crossing year as a single data set, regardless of parental genotypes.
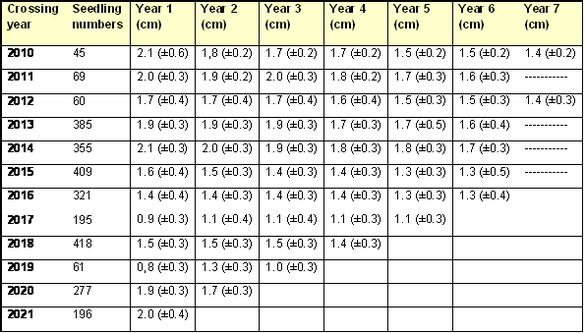
The results shown below are measurements to the end of the 2022 growing season, on seedlings from crosses made between 2010 and 2021. Mean values (± standard deviation) are given for seedling heights (Table 1), cumulative number of leaf nodes (Table 2), number of nodes produced annually (Table 3) and mean distance along the stem between successive nodes (Table 4).
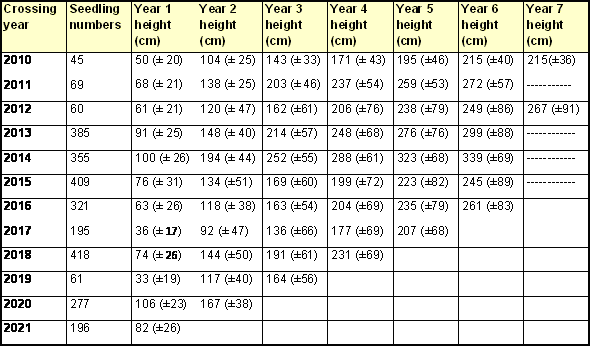
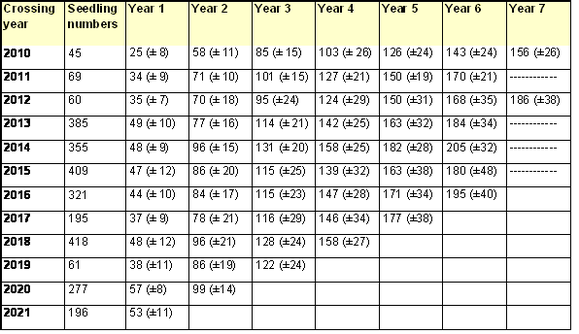

The results show a marked effect of crossing year on mean seedling heights, and on the number of nodes, attained after one year of growth. Whilst this variation is partly attributable to differences in weather patterns, significant improvements in cultural practice were also made between 2011 and 2015. Consequently, the growth rates of seedlings from crosses made from 2013 onwards reflect our general expectation, with average heights across a seedling population of 60-100 cm and 44-49 leaf nodes after the first year of growth, under good weather conditions. Similarly, we expect an average of at least 77 nodes by the end of the second year’s growth, and an average of at least 122 nodes by the end of the fourth year's growth.
The poor performance of seedlings from 2017 crosses during their first year of growth, with average seedling heights of 36 cm (Table 1) and just 37 leaf nodes (Table 2), was probably caused by the very dry growing conditions during the summer of 2018; our watering regime did not fully compensate for the lack of rainfall. The reasons for the equally poor first year performance of seedlings from 2019 crosses are unclear.
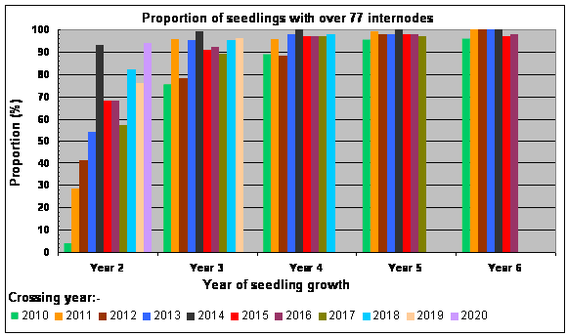
The increasing proportions of seedlings attaining 77 internodes (transition from the juvenile phase to the adult vegetative phase) after successive years of growth, between 2012 and the end of the 2022 growing season, are shown in the bar chart opposite.
Under our growing conditions none of the seedlings reach the adult vegetative phase after one year’s growth, but over 50% of seedlings from crosses made between 2013 and 2020 reached this phase after two years of growth. The general upward trend in the Year 2 proportions with successive crossing years between 2010 and 2020 probably reflects, among other factors, the improvements we've made in cultural practice over time.
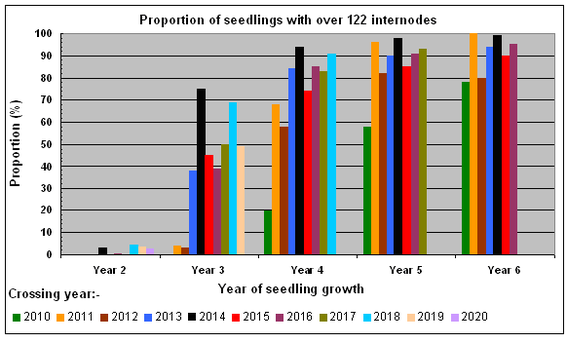
The increasing proportions of seedlings achieving >122 internodes (transition from adult vegetative to adult reproductive phase of development) after successive years of growth are shown in the bar chart opposite. Well over 50% of seedlings from crosses made between 2011 and 2018 achieved this transition after four years of growth. Seedlings from crosses made in 2014 performed very well, with 75% achieving >122 internodes after just three years of growth.
Frequency distributions for seedling height and number of internodes after two years of growth are shown opposite, for seedlings produced by crosses made in 2013.
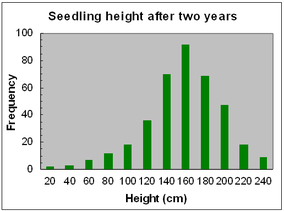
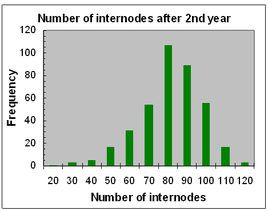
Both traits approximate to normal distributions, with wide variation around the mean, with heights ranging from 20-240 cm and number of internodes from 30-120.

The frequency distribution for numbers of internodes becomes flatter and more dispersed around the mean value during the first few years of growth. This trend is illustrated in the figure opposite for seedlings from crosses made in 2013, after one, two, three and four years of growth.

The relationship between seedling height and number of internodes for this data set is shown opposite. Linear regression of internode number on height showed a strong positive correlation between the variables (R2 = 0.79).
However, the spread of data around the line of best fit suggests that using height measurements to predict the number of internodes on the basis of regression equations is unlikely to be very accurate.
Effects of damage by thrips
Thrips (possibly Western Flower Thrip) caused serious damage to the terminal bud and expanding leaves on the main leader of many of our first year and second year seedlings during 2014 and 2015 (see photos below). Severe stunting of extension growth was accompanied by profuse, undesirable, lateral shoot production (right image), presumably triggered by the damage to the terminal bud and breaking of apical dominance. In some cases repeated, corrective, summer pruning was required to re-establish a leader. Damage has been progressively less severe between 2016 and 2019.



Seedlings from crosses made in 2013 were scored for thrip damage at the end of their second year of growth, classing them as 'severe', 'moderate' or 'undamaged'. Severe damage reduced both seedling height and number of internodes as shown in the figure opposite and the accompanying table below.
Differences between the means for severely damaged seedlings and those for moderate or undamaged seedlings were significant at p<0.001. The differences between moderate and undamaged seedlings were statistically insignificant.

Conclusions
Based on measurements to the end of the 2018 growing season, our conclusions regarding apple seedling growth under the prevailing environmental conditions in West Wales are:
-
Average seedlings heights of 90-100 cm with 44-49 nodes are attainable after one year of growth under good weather conditions. This compares
favourably with heights of 20-60 cm expected under good field conditions according to MAFF (1963). However, both seedling heights and number of nodes are reduced by poor weather during the
growing season, particularly by drought.
- Averages of 77 nodes or more are attainable after two years of growth.
- Averages of 122 nodes or more are attainable after four years of growth.
-
Whilst a few seedlings may flower as early as year three of growth, well over 50% of seedlings should definitely flower during year five;
these flowers having been initiated during the previous summer.
- The annual increments in seedling height and number of nodes decrease after the first year of growth. This underlines the importance of maximising seedling growth during the first year.
- The average distance between successive nodes is about 2 cm, so a rough estimate of the number of nodes on a seedling is given by dividing the height (cm) by two.
- Damage by thrips causes significant reductions in seedling height and the number of internodes attained after two years of growth. This is not easily controlled.
Seedling measurements will continue during 2020 and beyond, focusing on the transition from ‘adult vegetative phase’ to ‘adult reproductive phase,’ once 122 nodes
are achieved on the single leading shoot, and subsequent flowering rates. However, no further measurements will be made on seedlings from crosses made during 2010 and 2011, as the majority of
these seedlings have flowered and subsequently cut back to heights of 6-7 ft.
Reference
MAFF. 1963. Fruit Tree Raising: Rootstocks and Propagation Bulletin 135. HMSO London. 4th ed. 52p.