Pruning juvenile seedlings
Updated September 2021
Pruning affects the growth, shape and productivity of apple trees, whether young or old, grafted or grown from seed. Whilst many proven pruning systems have been developed, it remains the greatest single challenge to commercial growers aiming to maintain regular high yields of optimal quality fruit (Worraker and Withnail, 2013).
Seedling apple trees should not require pruning during the juvenile phase of growth. This is because seedling vigour and the duration of the juvenile period are inversely related (Visser, 1964). Consequently, any action restricting shoot growth lengthens rather than shortens the duration of the juvenile phase, and delays the onset of flowering (Janick et al. 1996). Most seedlings begin life with a single leading shoot. To encourage rapid and continuous extension growth it is advisable not to cut this shoot back until flowering and fruiting is achieved (Zimmerman, 1971). This advice makes sense given that the apical (youngest) shoot growth is the first to emerge from the juvenile phase (van Nocker and Gardiner, 2014). However, after the transition to the ‘adult vegetative phase’ (77 nodes), factors that reduce apical dominance of the leading shoot can promote lateral flower-bud formation (Hanke et al. 2007).
Pruning at Ystwyth Valley Apple Breeders

We grow our seedlings with a single leading shoot, and don’t prune this until the seedling flowers. The only exception to this rule is when the shoot is damaged, or stunted, by disease (e.g. canker) or pests (e.g. aphids and thrips). If this happens, we cut back the shoot to the youngest healthy leaf bud, whatever the time of year.
In contrast, we prune many of the lateral shoots arising from the single leader. This is necessary because our seedlings are grown closely spaced (23 cm apart) in pots (Fig. 1). Lateral shoots are usually produced during the first summer of growth, or early in the second year. Unchecked, they can soon lead to overcrowding and shading between adjacent seedlings. To prevent this, we prune laterals in late summer and again in winter. Viewing a row of seedlings running left to right (see Fig. 1), we cut off entirely most (75%) of the lateral shoots growing out forwards or backwards from the leading shoot. Lateral shoots growing out either side of the leader are pruned back to 3-5 leaves (summer) or 3-5 leaf buds (winter), if they are likely to interfere with adjacent seedlings. However, we leave the remainder unpruned, in case the seedling turns out to bear flowers on the tips of new shoots.
In terms of tree structure, our ultimate aim is to achieve a columnar growth form, with shortened laterals developing into fruiting spurs and dards (short laterals
<7-8 cm, ending in a fruiting bud), once the juvenile phase is over. In this respect the pruning regime is similar to that recommended for minarette trees. Some of the unpruned laterals should
develop into ‘brindles’. These are thin lateral shoots 8-30 cm in length which usually end in a fruit bud (Worraker and Withnail, 2013).
Pruning trial: effects of pruning lateral shoots

We began a small trial in 2016 to measure how pruning lateral shoots affects (a) the production of leaf nodes along the single leading shoot, and (b) seedling height. We expected that cutting back lateral shoots might promote extension growth of the leading shoot, due to the removal of competing sinks for water and nutrients. If so, this might increase the annual production of leaf nodes along the leader and shorten the juvenile phase.
Methods
The seedlings for the trial were grown from pips planted in January 2016, produced from crosses made in 2015. Following our usual procedure, they were transplanted into individual 5 litre pots during June 2016. The pots were partially sunk into the soil and arranged in closely spaced rows in a nursery bed outdoors (Fig.1). Seedlings from the same cross were adjacent to each other. The trial began on 5th August 2016, during the first year of seedling growth. Twenty-four matched pairs of seedlings were identified in a single nursery bed made up of four parallel rows of seedlings. To make a pair the two seedlings had to be (a) growing immediately adjacent to each other, (b) progeny from the same cross, and (c) of similar height and lateral shoot development, at the start of the trial.

The 24 pairs covered a wide range of starting heights and parental varieties. (Fig.3).
One seedling in each pair was designated as ‘unpruned’, and the other as ‘pruned’ (Fig. 2). The ‘unpruned’ treatment consisted of leaving the leading shoot and all lateral shoots unpruned for the duration of the trial. The ‘pruned’ treatment comprised no pruning of the leading shoot and our standard pruning regime for lateral shoots. The majority (75%) of lateral shoots growing out forwards or backwards from the leading shoot, when viewed from the front, were cut back flush with the leader. Half the lateral shoots growing out either side of the leader were pruned back to 3-5 leaves, or leaf buds. The rest were left unpruned. Seedlings were pruned in late summer and again during winter, when they were dormant.
Seedling height and the number of leaf nodes on the single leader were measured at the start of the trial and at the end of each successive year of growth. The statistical significance (at 95%) of differences in mean heights and numbers of nodes between pruned and unpruned treatments was assessed using paired t-tests. We plan to continue the trial for five years (to winter 2020/2021). The results will be updated here each year.
Results 2017 (Year 2)
Mean seedling heights and numbers of leaf nodes on the single leading shoot of 'unpruned' and 'pruned' seedlings are compared in the bar charts (Figs 4 & 5).
Mean values with standard deviations are also given in the summary table below.


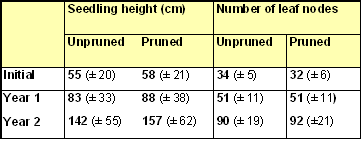
Mean seedling heights showed a small but statistically significant (p=0.002) difference between unpruned (55 cm) and pruned (58 cm) seedlings at the start of the trial. The difference in heights increased during Year 1, and further during Year 2. However, it was just statistically insignificant (p = 0.07) in both years. The high standard deviations for the Year 2 seedling heights reflect the wide variation in height between the different pairs of seedlings (Fig.6). Several pairs showed big differences in height between unpruned and pruned seedlings by the end of Year 2, although the differences remained modest for the majority of pairs.



The effect of pruning on the mean number of leaf nodes along the leading shoot was statistically insignificant, after Year 1 (p=0.85) and Year 2 (p=0.44) of growth. The results were remarkably similar after two years of growth for unpruned (90 nodes) and pruned (92 nodes) seedlings. Furthermore, with one or two exceptions, the differences in number of leaf nodes within pairs of seedlings remained modest after 2 years of growth (Fig. 7). The photograph opposite shows the differences in lateral shoots between pruned (LHS) and unpruned (RHS) members of a seedling pair typical after Year 2.
Conclusions (Year 2)
- Seedling height is not significantly influenced by pruning lateral shoots.
- The number of leaf nodes along the leading stem is not significantly affected by pruning lateral shoots.
- It is unlikely that the juvenile phase is significantly shortened by summer and winter pruning of lateral shoots.
Results 2018 (Year 3)
Measurements were limited to 20 seedling pairs due to the death of several seedlings from canker.


Mean seedling heights and numbers of leaf nodes on the single leading shoot of 'unpruned' and 'pruned' seedlings at the end of the 2018 growing season are compared in the bar charts (Figs 8 & 9) and summary table.
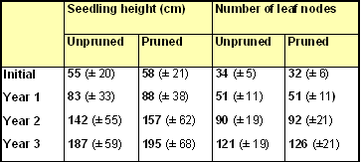
Mean seedling heights showed a small, but statistically insignificant (p=0.32),
difference between unpruned (187 cm) and pruned (195 cm) seedlings at the end of year 3. The high standard deviations for the Year 3 seedling heights reflect
the wide variation in height between the different pairs of seedlings (Fig.10). Pruned seedlings were tallest in twelve out of twenty pairs.


The mean number of leaf nodes along the leading shoot was higher in pruned seedlings (126 nodes) compared with unpruned seedlings (121 nodes) after Year 3 of growth, although the difference was statistically insignificant (p=0.12). Pruned seedlings had more nodes in eleven out of twenty pairs (Fig. 11).
Conclusions (Year 3)
- Seedling height is not significantly influenced by pruning lateral shoots.
- The number of leaf nodes along the leading stem is not significantly affected by pruning lateral shoots.
- It is unlikely that the juvenile phase is significantly shortened by summer and winter pruning of lateral shoots.
Results 2019 (Year 4)
Measurements were limited to 19 seedling pairs due to the death of a further seedling from canker.


Mean seedling heights and numbers of leaf nodes on the single leading shoot of 'unpruned' and 'pruned' seedlings at the end of the 2019 growing season are compared in the bar charts (Figs 12 & 13) and summary table.
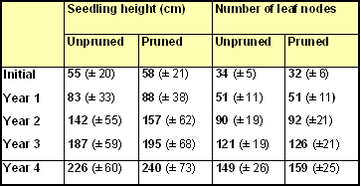
Mean seedling heights showed a statistically insignificant (p=0.30), difference
between unpruned (226 cm) and pruned (240 cm) seedlings at the end of year 4. The high standard deviations for the Year 4 seedling heights reflect the wide
variation in height between the different pairs of seedlings (Fig.14). Pruned seedlings were tallest in nine out of nineteen pairs after four years of the trial.

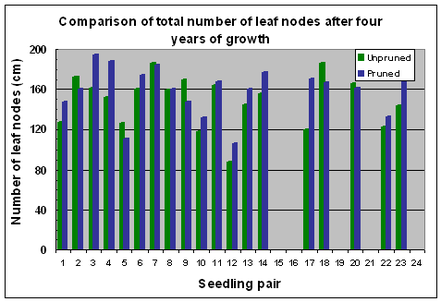
The mean number of leaf nodes along the leading shoot was higher in pruned seedlings (159 nodes) compared with unpruned seedlings (149 nodes) after Year 4 of growth, and the difference was statistically significant (p=0.04). Pruned seedlings had more nodes in thirteen out of nineteen pairs (Fig. 15).
Conclusions (Year 4)
- Seedling height is not significantly influenced by pruning lateral shoots.
- The number of leaf nodes along the leading stem is significantly higher in seedlings subject to pruning lateral shoots.
- It is unlikely that the juvenile phase is significantly shortened by summer and winter pruning of lateral shoots.
Results 2020 (Year 5)
Measurements were limited to 18 seedling pairs due to the death of a further seedling from canker.


Mean seedling heights and numbers of leaf nodes on the single leading shoot of 'unpruned' and 'pruned' seedlings at the end of the 2020 growing season (year 5) are compared in the bar charts (Figs 16 & 17) and summary table.

Mean seedling heights showed a statistically insignificant (p=0.58), difference between unpruned (259 cm) and pruned (267 cm) seedlings at the end of year 5. The high standard deviations for the Year 5 seedling heights reflect the wide variation in height between the different pairs of seedlings (Fig.18). Pruned seedlings were tallest in ten out of eighteen pairs after five years of the trial.


The mean number of leaf nodes along the leading shoot was higher in pruned seedlings (189 nodes) compared with unpruned seedlings (178 nodes) after Year 5 of growth, and the difference was statistically insignificant (p=0.11). Pruned seedlings had more nodes in thirteen out of eighteen pairs (Fig. 19).
Five seedlings subsequently flowered in 2021 (Year 6) in both pruned and unpruned treatments.
Conclusions (Year 5)
- Seedling height is not significantly influenced by pruning lateral shoots.
- The number of leaf nodes along the leading stem is not affected by pruning lateral shoots.
- It is unlikely that the juvenile phase is significantly shortened by summer and winter pruning of lateral shoots.
References
Hanke M-V, Flachowsky H, Peil A, Hättasch C. 2007. No Flower no Fruit – Genetic Potentials to Trigger Flowering in Fruit Trees. Genes, Genomes and Genomics 1(1), 1-20.
Janick J, Cummins JN, Brown SK and Hemmat M. 1996. Apples. In: Janick J, Moore JN, eds, Fruit Breeding, Volume I: Tree and Tropical Fruits. John Wiley & Sons, Inc. 1-77.
Van Nocker S and Gardiner SE. 2014. Breeding better cultivars, faster: applications of new technologies for the rapid deployment of superior horticultural tree crops/ Horticultural Research 1, 14022; doi:10.1038/hortres.2014.22.
Visser T. 1964 Juvenile phase and growth of apple and pear seedlings. Euphytica 13, 119-129.
Worraker R and Withnail M . 2013. The Apple Pruning Manual. Esquire Print, Nettlestead, Kent, UK. 117p. ISBN No. 978-0-9531850-1-6.
Zimmerman RH. 1971. Flowering in crab apple seedlings: methods of shortening the juvenile phase. Journal of American Society of Horticultural Science 96, 404-411.